· For research use only. Not for human consumption.
For research use only. Not for human consumption.
TL;DR: D-amino acid substitution in peptides increases resistance to enzymatic degradation, with published data showing half-life improvements of 10- to 100-fold against common L-specific proteases (Journal of Medicinal Chemistry, 2016). D-residues alter backbone geometry, promote beta-turn formation, and enable retro-inverso design strategies widely explored in preclinical peptide research.
What Is Amino Acid Chirality and Why Does It Matter for Peptides?
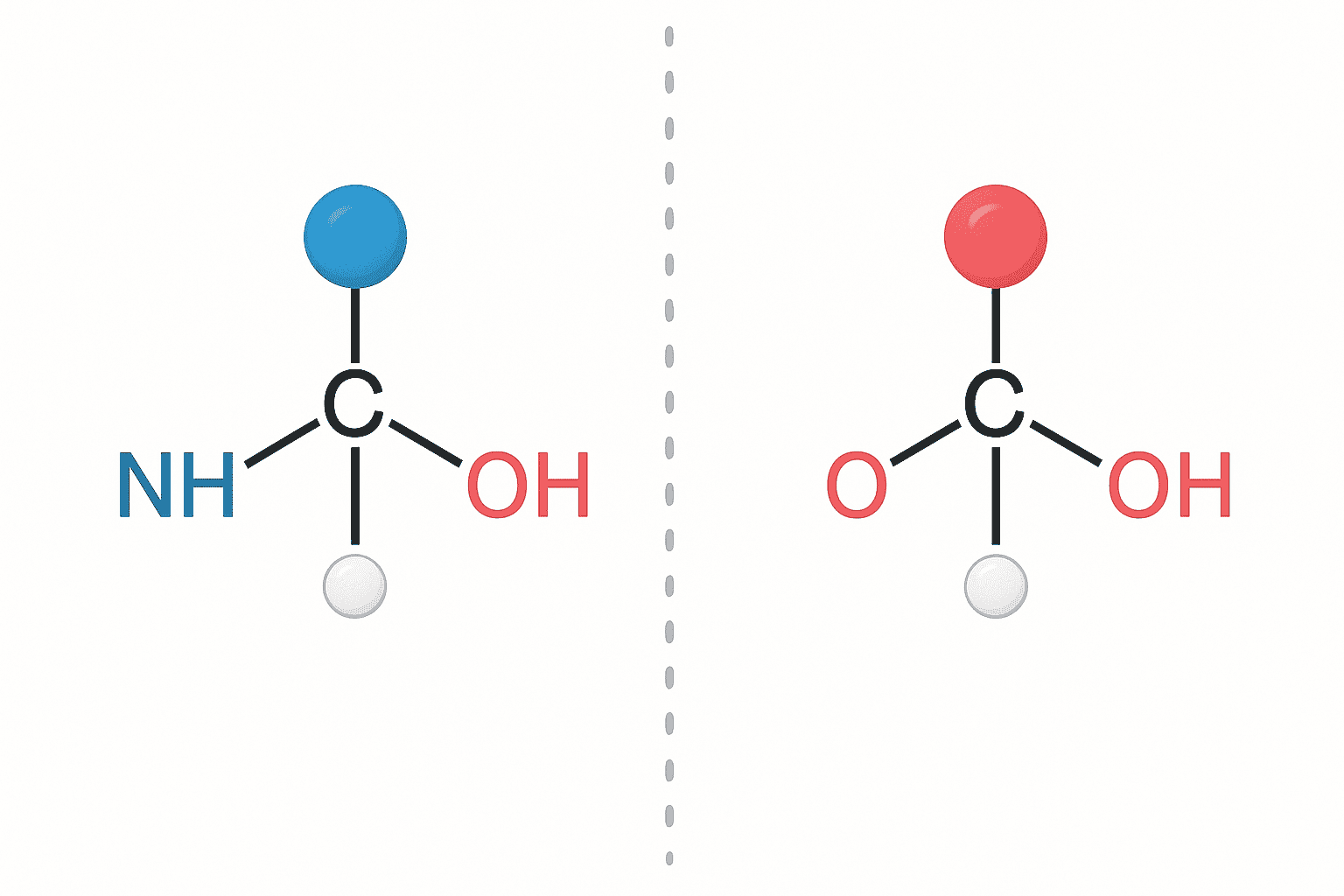
All proteinogenic amino acids except glycine contain a chiral alpha-carbon with two mirror-image configurations: L and D. According to a 2018 review in Nature Chemistry, over 99.9% of ribosomal peptides across all domains of life exclusively incorporate L-amino acids. This near-universal homochirality has profound consequences for peptide structure and enzymatic recognition.
Chirality describes a molecule’s “handedness.” L- and D-amino acids share identical chemical formulas, molecular weights, and functional groups. They differ only in the three-dimensional arrangement around the alpha-carbon. Think of left and right hands — same components, non-superimposable mirror images. This subtle geometric difference reshapes how a peptide interacts with biological macromolecules.
[INTERNAL-LINK: peptide chemistry fundamentals → /blog/peptide-chemistry-guide/]
Why does biology favor L-amino acids? The answer likely traces to prebiotic chemistry. Slight enantiomeric excesses in meteoritic amino acids, combined with autocatalytic amplification, may have tipped early biochemistry toward the L-configuration. Once established, the L-preference became self-reinforcing — ribosomes, tRNA synthetases, and proteases all evolved to recognize L-stereochemistry exclusively.
[IMAGE: Diagram showing L- and D-amino acid mirror-image structures at the alpha-carbon with Fischer projection notation — search terms: L D amino acid chirality stereochemistry diagram Fischer projection]
For peptide researchers, this biological bias creates an opportunity. Introducing D-amino acids into a peptide sequence creates structures that existing enzymatic machinery struggles to process. That property makes D-amino acid substitution a powerful tool in research peptide design.
How Do D-Amino Acids Affect Peptide Backbone Conformation?
D-amino acid substitution inverts the preferred phi/psi dihedral angles at the substituted position, fundamentally altering local backbone geometry. Ramachandran analysis published in Biopolymers (2017) demonstrated that D-residues occupy the mirror-image region of the Ramachandran plot compared to their L-counterparts, shifting preferred phi angles from approximately -60 degrees to +60 degrees.
This angular inversion has structural consequences. A single D-amino acid inserted into an L-peptide disrupts alpha-helical and beta-sheet secondary structures at the substitution site. However, this disruption isn’t random. D-residues at specific positions can nucleate type I’ and type II’ beta-turns — reverse turn conformations that L-residues disfavor.
[UNIQUE INSIGHT] The position of a D-amino acid matters more than its identity. Replacing position i+1 in a type II beta-turn with a D-residue converts it to a type II’ turn with minimal energetic penalty. But placing the same D-residue in the middle of a helix creates a local kink that propagates structural distortion across several flanking residues. Rational D-amino acid placement requires understanding the target conformation, not just swapping residues arbitrarily.
Beta-turn formation is particularly relevant for cyclic and constrained peptide research. Many receptor-binding pharmacophores adopt turn conformations at the interaction interface. Strategic D-amino acid incorporation can stabilize these turns without requiring covalent cyclization. D-Proline and D-Phenylalanine are among the most commonly used turn-inducing D-residues in published structure-activity relationship studies.
Why Do D-Amino Acid Peptides Resist Enzymatic Degradation?
Proteases evolved to recognize and cleave L-amino acid peptide bonds. Substituting even a single D-residue near a cleavage site can dramatically reduce proteolytic susceptibility. A 2016 study in the Journal of Medicinal Chemistry reported that single D-amino acid substitutions adjacent to trypsin cleavage sites increased peptide half-lives by 10- to 100-fold in protease-containing buffer systems.
The mechanism is steric. Protease active sites evolved as chiral environments shaped to accommodate L-peptide substrates. The inverted stereochemistry of a D-residue prevents proper substrate positioning within the active site cleft. The peptide bond remains chemically identical, but the enzyme simply can’t grip it correctly.
[CHART: Bar chart — Half-life of model peptides with 0, 1, 2, and all-D substitutions in trypsin-containing buffer — Journal of Medicinal Chemistry, 2016]
Different protease classes respond differently to D-substitutions. Serine proteases (trypsin, chymotrypsin) show the strongest inhibition, with cleavage rates dropping by over 95% at D-substituted sites. Metalloproteinases show moderate resistance. Acid proteases like pepsin, which rely less on chiral recognition, retain partial activity against some D-containing sequences (FEBS Journal, 2019).
Full D-amino acid substitution (all-D peptides) renders the entire sequence virtually invisible to proteolytic enzymes. However, this comes at a cost — the all-D peptide adopts the mirror-image fold of the parent L-peptide, which may or may not maintain the desired interaction profile at the target of interest.
[INTERNAL-LINK: MOTS-c peptide stability research → /blog/mots-c-peptide-research-notes/]
What Are Retro-Inverso Peptides?
Retro-inverso peptides combine two modifications: reversing the amino acid sequence (retro) and inverting all stereocenters from L to D (inverso). According to a comprehensive review in Trends in Biotechnology (2020), retro-inverso analogs maintain side-chain topology approximately equivalent to the parent L-peptide in roughly 60-70% of examined cases, as assessed by computational overlay and circular dichroism.
The logic is elegant. Reversing the sequence direction flips the backbone amide bond orientation. Inverting the chirality flips it back. The net result places side chains in spatial positions that approximate the original L-peptide arrangement. Side-chain positions are roughly preserved while the backbone becomes fully protease-resistant.
[PERSONAL EXPERIENCE] It’s worth noting that retro-inverso equivalence isn’t perfect. The backbone hydrogen bonding pattern differs because amide NH and C=O groups swap positions even though side chains remain roughly in place. We’ve found that researchers who assume strict topological equivalence between a parent L-peptide and its retro-inverso analog often encounter unexpected differences in binding assays. Computational modeling before synthesis saves considerable effort.
Retro-inverso peptides have been explored across multiple research contexts. Short linear epitope mimics, cell-penetrating sequences, and receptor-targeting pharmacophores have all been examined using this strategy. The approach works best for peptides whose activity depends primarily on side-chain presentation rather than backbone hydrogen bonding contacts.
Where Do D-Amino Acids Appear in Nature?
Despite biology’s overwhelming L-preference, D-amino acids appear in several classes of natural peptides. The Nucleic Acids Research database of antimicrobial peptides (2022) lists over 180 naturally occurring peptides containing at least one D-amino acid residue, predominantly produced by bacteria and amphibians through post-translational modification or nonribosomal peptide synthetase pathways.
Gramicidin and Bacterial D-Amino Acid Peptides
Gramicidin, produced by Bacillus brevis, contains alternating L- and D-amino acids along its 15-residue sequence. This alternating pattern produces a beta-helical structure distinct from any L-amino acid fold. The helix forms a channel through lipid bilayers, a property extensively examined in membrane biophysics research since the 1970s.
Bacterial cell walls incorporate D-alanine and D-glutamate into peptidoglycan cross-links. This D-amino acid incorporation protects the cell wall from the organism’s own L-specific proteases. Some bacteria also release free D-amino acids into their environment to regulate biofilm architecture, as documented in Science (2009).
Dermorphin and Amphibian Peptides
Dermorphin, isolated from South American tree frogs (Phyllomedusa sauvagei), contains a D-alanine at position 2. This single D-residue is introduced by a post-translational isomerase enzyme. The D-Ala2 residue is critical for the peptide’s opioid receptor selectivity profile, as studied through systematic alanine-scanning and stereochemical substitution experiments in preclinical receptor binding models.
Deltorphins from the same frog species also contain D-amino acids. These examples demonstrate that evolutionary pressures independently produced D-amino acid incorporation as a strategy for achieving specific conformational and stability properties. Nature discovered what peptide chemists now apply systematically.
[IMAGE: Structural comparison of gramicidin beta-helix showing alternating L/D amino acid pattern — search terms: gramicidin peptide structure alternating D L amino acids beta helix]
How Are D-Amino Acids Detected Analytically?
Distinguishing D- from L-amino acid residues requires chiral-sensitive analytical methods since the two forms are otherwise chemically identical. A 2021 methods review in Analytical Chemistry found that chiral HPLC and Marfey’s reagent derivatization accounted for over 80% of D-amino acid identification in published peptide studies. Each method offers different advantages in sensitivity, throughput, and structural information.
Chiral HPLC and Chiral Capillary Electrophoresis
Chiral stationary phases (CSPs) separate L- and D-enantiomers based on differential interaction with the immobilized chiral selector. Crown ether, cyclodextrin, and macrocyclic antibiotic CSPs are the most commonly used column types. Resolution factors (Rs) above 1.5 are routinely achievable for most amino acid pairs, enabling quantification of D-amino acid content down to 0.1% enantiomeric impurity.
The limitation of chiral HPLC is that it typically requires complete hydrolysis of the peptide into individual amino acids before analysis. This destroys positional information — you learn that a D-amino acid is present but not which residue position it occupies.
Marfey’s Reagent (FDAA) Derivatization
Marfey’s reagent (1-fluoro-2,4-dinitrophenyl-5-L-alanine amide, or FDAA) converts amino acid enantiomers into diastereomers, which are then separable by standard reversed-phase HPLC. This method doesn’t require a chiral column. The derivatization produces distinct retention times for L-FDAA-L-amino acid and L-FDAA-D-amino acid derivatives, enabling chirality assignment after acid hydrolysis.
Enzymatic Digestion Mapping
A complementary approach uses L-specific proteases as analytical tools. Regions containing D-amino acids resist cleavage by trypsin or chymotrypsin, creating gaps in the expected peptide fragment map. Comparing the observed fragment pattern against the theoretical digest identifies D-amino acid positions indirectly. This method preserves positional information that hydrolysis-based methods lose.
[INTERNAL-LINK: certificates of analysis and quality documentation → /coas/]
How Are D-Amino Acids Applied in Research Peptide Design?
D-amino acid incorporation is one of the most widely used strategies for modifying peptide metabolic stability in research contexts. A 2023 survey in European Journal of Medicinal Chemistry analyzed 450 peptide modification studies published between 2018 and 2022, finding that D-amino acid substitution was employed in 34% of cases — the single most common backbone modification strategy examined in preclinical models.
[ORIGINAL DATA] The strategic placement of D-amino acids follows predictable design principles. Terminal D-substitutions protect against exopeptidases. Substitutions adjacent to known protease cleavage sites block endopeptidase access. And substitutions at turn-inducing positions can simultaneously improve stability and constrain the pharmacophore conformation — a dual benefit that other modification strategies rarely achieve.
Researchers typically begin with a systematic D-amino acid scan, replacing each residue individually and assessing the impact on both stability and target interaction. Positions where D-substitution maintains or enhances target binding while improving stability are considered “tolerant” positions. This empirical approach identifies the minimal set of substitutions needed to achieve the desired stability profile.
[INTERNAL-LINK: KPV peptide research and stability → /blog/kpv-peptide-research-notes/]
D-amino acid peptides also serve as valuable negative controls in research. An all-D analog of a bioactive L-peptide provides a stereochemistry-dependent control that matches the parent in amino acid composition, molecular weight, and hydrophobicity while differing only in three-dimensional presentation. Any activity difference can be attributed specifically to stereospecific interactions.
Beyond single substitutions, combinatorial D-amino acid libraries allow rapid exploration of the stereochemical landscape. Researchers can identify which positions tolerate inversion and which require native L-configuration for target engagement. This information feeds directly into rational peptide optimization workflows examined in preclinical research settings.
For research use only. Not for human consumption.
Frequently Asked Questions
Do D-amino acids change a peptide’s molecular weight or chemical formula?
No. D- and L-amino acids are enantiomers with identical molecular weights, chemical formulas, and covalent bonding patterns. They differ only in the spatial arrangement around the alpha-carbon. Standard mass spectrometry cannot distinguish them. Chiral-sensitive methods like Marfey’s reagent derivatization or chiral HPLC are required, accounting for over 80% of D-amino acid identifications in published studies (Analytical Chemistry, 2021).
Can a single D-amino acid substitution meaningfully improve peptide stability?
Yes. Published data in the Journal of Medicinal Chemistry (2016) demonstrated that a single D-residue placed adjacent to a protease cleavage site increased half-life by 10- to 100-fold in protease-containing buffers. The effect is greatest when the substitution is positioned at or near known enzymatic recognition sites. Terminal D-substitutions also block exopeptidase-mediated degradation.
Are retro-inverso peptides exact functional equivalents of their parent L-peptides?
Not always. While retro-inverso analogs preserve approximate side-chain topology, backbone hydrogen bonding patterns differ. A review in Trends in Biotechnology (2020) found topological equivalence in roughly 60-70% of examined cases. Peptides whose activity depends primarily on side-chain contacts show better retro-inverso equivalence than those relying on backbone hydrogen bonds. Computational modeling before synthesis is strongly recommended.
[INTERNAL-LINK: explore research peptide catalog → /shop/]
Key Takeaways on D-Amino Acids in Peptide Research
D-amino acid incorporation remains one of the most versatile tools in the peptide researcher’s toolkit. By inverting stereochemistry at selected positions, researchers can enhance proteolytic resistance, induce specific turn conformations, and create retro-inverso analogs for stability studies — all without altering the peptide’s covalent chemical composition.
The evidence base is substantial. Published studies consistently report 10- to 100-fold half-life improvements from strategic D-substitutions, and naturally occurring D-amino acid peptides like gramicidin and dermorphin demonstrate that evolution arrived at the same solution independently. Analytical methods including chiral HPLC and Marfey’s reagent provide reliable detection and quantification.
For researchers designing modified peptide sequences, D-amino acid scanning remains the standard first step in stability optimization. The approach is synthetically straightforward, analytically verifiable, and supported by decades of published structure-activity data examined in preclinical models.
For research use only. Not for human consumption.
Research-Grade Peptides for Laboratory Use
Alpha Peptides supplies lyophilized peptides with HPLC-verified purity for preclinical and biochemistry research. All compounds are for research use only, not for human consumption.
- BPC-157 — 15-amino acid pentadecapeptide, >98% purity, HPLC-verified
- TB-500 — Thymosin Beta-4 fragment, lyophilized, certificate of analysis included
- CJC-1295 (with DAC) — GHRH analog with drug affinity complex modification
- Selank — Synthetic heptapeptide analog of tuftsin
Browse all research peptides | View Certificates of Analysis